Data on the resting metabolism of the body and the central nervous system were sought in the existing literature. The search was carried out as systematically as possible through use of the Science Citation Index and bibliographies of relevant papers and books, Resting and basal metabolism of the body was found to be available for a large number of individuals of various weights and species (8, 13). Metabolism of the entire nervous system, on the other hand, has not been determined directly ; therefore all calculations of neural metabolism were made by multiplying metabolic rates of brain and spinal cord tissue by the corresponding tissue weight for each species. The obtained values for brain and spinal cord metabolism were then summed to yield the total resting metabolism of the central nervous system.
Brain weight data were found to be available for many vertebrate species. A number of investigators have documented the relationship between the size of the brain and the size of the body of individual vertebrates (12, 20),
Spinal cord weights were available for fewer vertebrates than were brain weights. Several investigators have collected spinal cord weight data for a number of vertebrate species, They include Latimer and co-workers (56-61) and Krompecher and Lipák (52). Direct data were available for the dolphin (79), whereas for the whale and alligator, spinal cord weights could be estimated from planimetric measurements and extrapolations from data in the literature (77, 89).
We determined the spinal cord weights of four cold- blooded vertebrates from our own specimens. The entire central nervous system was dissected from formaldehyde- fixed specimens of the dogfish, shark, perch, bullfrog, and goldfish after the entire specimen had been weighed. The brain and spinal cord were severed at their junction and the dura and spinal and cranial nerves were removed. The ratio of spinal cord to brain weight was computed. This ratio was subsequently applied to published brain weight data of other individuals of the same species, in order to estimate corresponding spinal cord weights.
In some species for which spinal cord weights were not available, we used spinal cord-to-brain weight ratios from other related species with similar body shape and size. The following extrapolations were made from one species to another: from frog to toad; from tree squirrel to ground squirrel; from guinea pig to opossum; from dog to fox; from brown bat to vampire bat; from macaque to baboon, from emu to ostrich; from human to chimpanzee; from sheep to pig; and from horse to camel.
Direct measurements of in vivo rates of oxygen consumption of the whole brain were found in the literature for five species and have been plotted in Fig. I. The data were obtained for the rat (68), cat (30), dog (32), rhesus monkey (84), and human (50). A regression equation was fitted to these data and took the form of
log y = 0.86 - 0.13 log x (1)where y is brain metabolic rate in cm3 O2 · 100 g-1 · min -1 and x is brain weight in grams. The declining rates of metabolism in larger brains presumably reflect the lower packing densities of neuronal cell bodies, which is where most metabolism occurs (90).
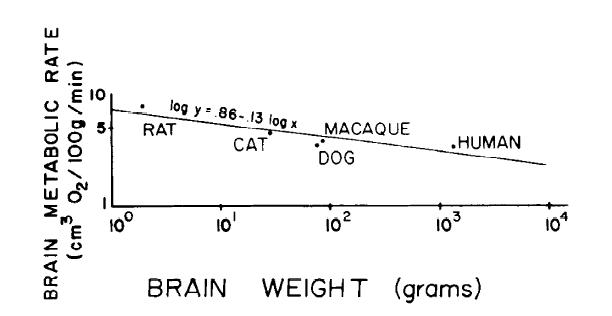
FIG. 1. Brain metabolic rate as a function of brain weight in warm-blooded vertebrates, Data were obtained in vivo in rat (681, cat (301, dog (32), rhesus monkey (84), and human (50). Data are represented on logarithmic axes and are fitted with a linear regression equation of log y = 0.86 - 0.13 log x (y, brain metabolic rate in cm3 O2 · 100 g-1 · min-1 ; x brain weight in grams.)
Brain metabolism of cold-blooded animals could only be estimated indirectly, because there are no in vivo determinations in the literature. Therefore, we have used the Q10 value for changes in nervous tissue metabolism as a function of temperature to calculate a function for nervous tissue at 20°C, the temperature used for most determinations of cold-blooded vertebrate metabolism. The value of Q10 was set at 2.1, which is the mean of empirical calculations for brain tissue by a number of different authors using both warm-blooded and cold-blooded vertebrates (29, 67, 71, 88). Setting Q10 equal to 2.1 and solving the following equation for the temperature coefficient of chemical reactions
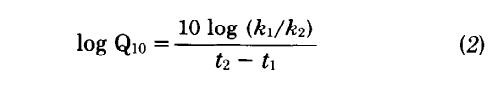
with t2 = 37°C and t1 = 2O°C, one obtains a ratio of k1/k2 equal to 3.5. This ratio is then used to convert Eq. 1 to a second equation for cold-blooded vertebrates in which each value is l/3.5 times the corresponding value for a warm-blooded vertebrate
log y = 0.32 - 0.13 log x (3)where y is the brain metabolic rate in cm3 O2 · 100 g-1 · min -1 and x is the brain weight in grams.
Spinal cord metabolic rates were found for only three species. Rosenberg (82) found that the spinal cord metabolic rate was 90% that of the brain in the European frog. McIlwain (66) found that guinea pig spinal cord slices respired at a rate 40% that of cerebral cortex slices. Hertz and Clausen (37) reported that slices of calf spinal cord respired at a rate 18% that of cerebral cortex slices. The latter two figures must be adjusted because cortex respiration is 40% greater than whole-brain respiration (23, 25, 66). Therefore, one may assume that guinea pig spinal cord metabolic rate is 56% that of whole brain and that the metabolic rate of calf spinal cord is 25% that of whole brain. These data, along with data from the frog, are plotted in Fig. 2 as a function of spinal cord weight and fitted with a regression equation
log y - log x = -0.25 - 0.16 log SW (4)in which y represents spinal cord metabolic rate in cm3 O2 · 100 g-1 · min-1 , x represents brain metabolic rate in the same terms, and SW represents the spinal cord weight in grams.
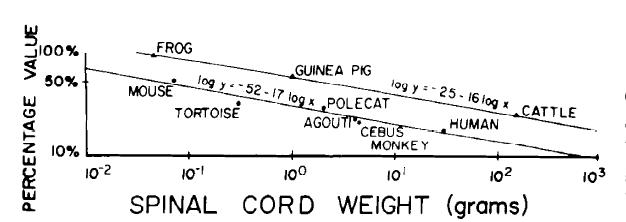
FIG, 2, Spinal cord metabolic rate as a percentage of brain metabolic rate (upper line) and volume of gray matter at cervical level as percentage of total spinal cord volume (lower line). Metabolic data are from published measurements of european frog (82), guinea pig (65), and calf (37). Equation of line fitted to these data takes form log (y/x) = - 0.25 - 0.16 log SW (y, spinal cord metabolic rate; x, brain metabolic rate; SW, spinal cord weight in grams). Data on spinal cord gray matter are from published measurements for mouse, Mus musculus (39), turtle, Testudo sp. (39), polecat, Putorius spa. (39), agouti rodent, Dasyprocta agouti (39), cebus monkey, Cebus sp. (39), and human (55). Slope of regression line for these data on logarithmic axes is -0.17, ▲, % value: spinal cord metabolic rate divided by brain metabolic rate. ●, 5% value: gray matter volume divided by total spinal cord volume.
Although basal metabolism data were available in the literature for all species under consideration, in some cases it was necessary to extrapolate from data in the literature to match the size of an individual for which brain and spinal cord weights were available. This was necessary because both basal metabolism and brain- to-body weight ratios are a function of the size of the individual animals. In those cases in which the body weights of animals for which the two sets of data were available differed by less than I0%, no adjustment was made. In case of differences greater than l0%, the metabolic rate was adjusted according to the following equation
log BM2 - log BM1 = 0.75 (log BW2 - log BW1) (5)where BM is body metabolism and BW is body weight (8)
The spinal cord metabolism of the elephant was treated as a special case. No data were available on the size of the spinal cord of a mature elephant. However, judging from the data on other large mammals, one could assume that the overall metabolism of the spinal cord is no more than 10% of the brain metabolism. For example, in the camel it is 9%, in cattle l0%, and in the horse 9%. Therefore, we used a figure of 9% of brain metabolism as an estimate for metabolism of the elephant spinal cord.
When data were available from many individuals of a
species, we chose data from mature, healthy individuals
as reported in the literature. A second consideration was
made in choosing which values to use. Insofar as possible,
we used animals of the same body weight for both the
body metabolism data and the brain and spinal cord
data.
 |
 |
 |